
Researchers
 DELANOUE Renald - +33 489150763FRANçOIS Charlotte - +33 489150763TOURNIéRE Océane - +33 489150763TOURNIéRE Océane - +33 489150763MESCHI Eleonora - +33 489150763
DELANOUE Renald - +33 489150763FRANçOIS Charlotte - +33 489150763TOURNIéRE Océane - +33 489150763TOURNIéRE Océane - +33 489150763MESCHI Eleonora - +33 489150763
PreDocs
LAVOCAT Laura - +33 489150763
Students
LETAPISSIER Maissane - +33 RPARVATI Ananya - +33 R
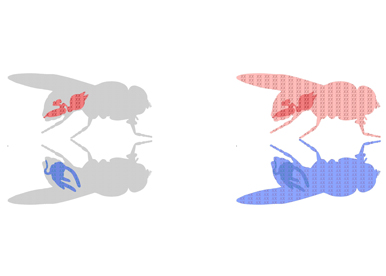
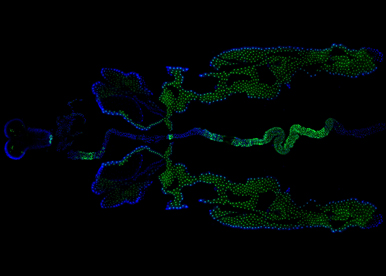
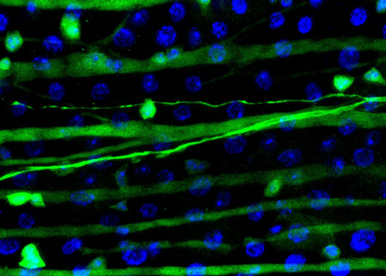
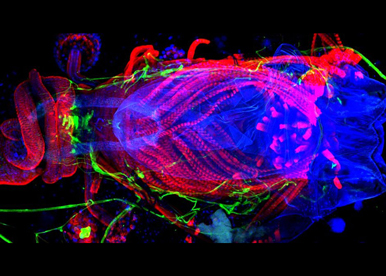
An Assistant Engineer position is available in the group “Cellular Sex and Physiology” at the Institute of Biology Valrose (iBV) in Nice, France. Our research focuses on developmental, metabolic, and behavioural sex differences, with the goal of understanding how sex chromosome constitution (XX versus XY) influences physiology across the body.
We are currently uncovering new genes, pathways, and conceptual frameworks driving sex chromosome effects, using multi-scale approaches that integrate biochemistry, genetics, and cell biology. Our model system is the fruit fly Drosophila melanogaster (see Hudry et al., 2019 Cell; Delanoue et al., 2023 Nat Ecol Evol; François et al., 2023 Nat Commun; Hérault et al., 2024 Nat Commun). More information is available on our lab webpage (http://ibv.unice.fr/research-team/hudry/).
This position is funded by an ERC Consolidator Grant for 3 years. We seek a highly motivated candidate with a strong interest in evolution and developmental biology. The work will involve biochemistry and molecular biology experiments, as well as embryo microinjection for Drosophila transgenesis. Team spirit and a positive, collaborative attitude are highly valued in our group.
The Institute of Biology Valrose (27 research teams, ~300 people, 25 nationalities) offers an international and dynamic scientific environment. English is the working language. The institute provides state-of-the-art core facilities and fosters a collaborative and stimulating atmosphere in a beautiful city and region.
If you are interested, please contact Bruno Hudry (Bruno.Hudry@univ-cotedazur.fr) with your CV, a brief description of your expertise, and up to three recommendation contacts.
A fully funded postdoc position is available in the Cellular Sex and Physiology group at the Institute of Biology Valrose (iBV) in Nice, France. Our research explores how sex chromosome composition (XX vs XY) influences physiology across different organs and systems, with a particular focus on developmental, metabolic, and behavioural sex differences. Our lab combines genetics, biochemistry, and cell biology in a multi-scale approach to uncover new genes, pathways, and fundamental principles that mediate sex chromosome effects. We use Drosophila melanogaster as a model system, with recent publications in Nature Communications (Hérault et al., 2024, François et al., 2023), Nature Ecology & Evolution (Delanoue et al., 2023), and Cell (Hudry et al., 2019). For more details, please visit our lab website (http://ibv.unice.fr/research-team/hudry/).
This position is funded by an ERC Consolidator Grant focused on X chromosome dosage compensation and is expected to start in 2025. We are looking for a highly motivated candidate with a strong interest in cell and developmental biology. Experience with live imaging and image analysis is essential. Prior experience with Drosophila is appreciated but not required. We value scientific curiosity, collaborative spirit, and a positive working environment.
The Institute of Biology Valrose (iBV) hosts 28 research teams and over 300 scientists from 25 nationalities. English is the working language, and the institute offers access to cutting-edge core facilities in a collaborative, dynamic, and supportive scientific community—located in the beautiful city of Nice on the French Riviera.
To apply, please contact Bruno Hudry (Bruno.Hudry@univ-cotedazur.fr) with the following: -a CV, -a brief statement of research interests and key skills and contact information for up to three referees.
A fully funded PhD position is available in the Cellular Sex and Physiology group at the Institute of Biology Valrose (iBV) in Nice, France. Our research explores how sex chromosome composition (XX vs XY) influences physiology across different organs and systems, with a particular focus on developmental, metabolic, and behavioural sex differences. Our lab combines genetics, biochemistry, and cell biology in a multi-scale approach to uncover new genes, pathways, and fundamental principles that mediate sex chromosome effects. We use Drosophila melanogaster as a model system, with recent publications in Nature Communications (Hérault et al., 2024, François et al., 2023), Nature Ecology & Evolution (Delanoue et al., 2023), and Cell (Hudry et al., 2019). For more details, please visit our lab website (http://ibv.unice.fr/research-team/hudry/).
This position is funded by an ERC Consolidator Grant focused on X chromosome dosage compensation and is expected to start in 2025. We are looking for a highly motivated candidate with a strong interest in cell and developmental biology. Experience with live imaging and image analysis is essential. Prior experience with Drosophila is appreciated but not required. We value scientific curiosity, collaborative spirit, and a positive working environment.
The Institute of Biology Valrose (iBV) hosts 28 research teams and over 300 scientists from 25 nationalities. English is the working language, and the institute offers access to cutting-edge core facilities in a collaborative, dynamic, and supportive scientific community—located in the beautiful city of Nice on the French Riviera.
To apply, please contact Bruno Hudry (Bruno.Hudry@univ-cotedazur.fr) with the following: -a CV, -a brief statement of research interests and key skills and contact information for up to three referees.

When the X Chromosome Makes the Difference
Read More

A young iBV researcher honored with the 2025 L’Oréal-UNESCO For Women in Science Young Talents France Award !
Read More

Bruno Hudry awarded an ERC Consolidator Grant 2024!
Read More

Bruno Hudry receives the CNRS Bronze Medal!
Read More

Bruno HUDRY nommé EMBO Young Investigator
Read More

UCA Annual Award Ceremony 2019: 4 iBV members recognized this year !
Read More
Intestinal sex differences in sugar metabolism regulates sperm production
Read More